

This is the fourth in a series of five blogs where I’ll discuss how metabolomics is set to transform medicine as we know it. The applications of omics in biomedical research are vast. To help organize the different ways metabolomics can be used, I’ll discuss this in the context of 5P medicine.
If you are already familiar with the concept of 5P medicine and how omics contribute to the transformation of medicine, click to move forward to the population-based medicine section of this blog.
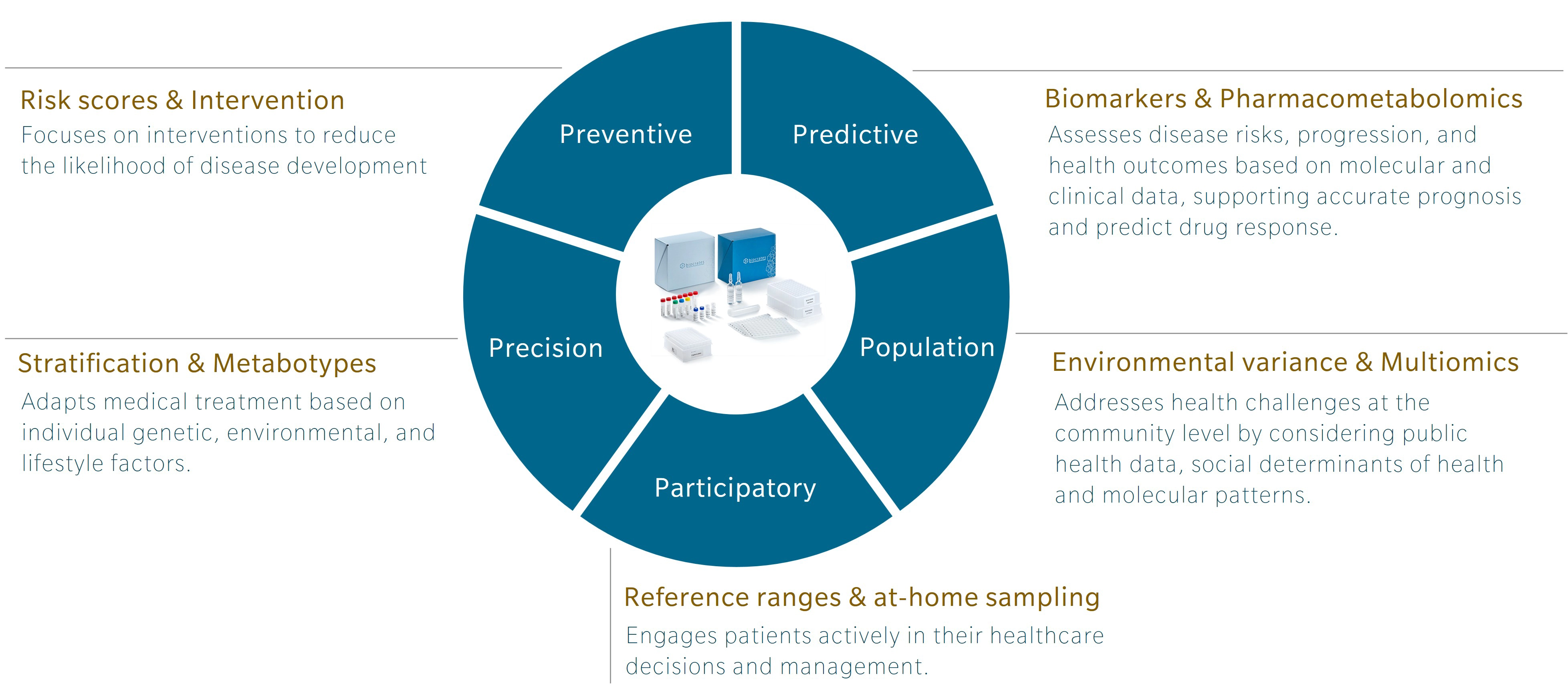
An introduction to 5P medicine
5P medicine is a concept developed to address the limitations of traditional Western medicine, which typically focuses on reacting to illness or injury. Making use of five components – preventive, predictive, precision, participatory and population-based medicine – 5P medicine aims to shift the focus towards a more proactive and patient-centric practice.
Personally, I first encountered this concept in a book by Leroy Hood and Nathan Price, The Age of Scientific Wellness. Hood and Price describe what they call P4 medicine. Rather than the familiar model that treats or manages disease after its occurrence, the authors offer an alternative that leverages the scientific tools at our disposal to understand health and disease. The result is a transition from what they call a “sickcare” system towards a genuine “healthcare” system. The 5P model expands this concept by adding population-based medicine to the original four and incorporating strategies that use the power of large cohort studies to find additional insights.
Omics and the future of research and health
Omics research has been around for over 30 years, and while genomics is gaining traction and beginning to be used in the clinics, other omics, and multiomics integration, are still lagging. Each omic addresses a distinct layer of biology, with its own codes, regulatory signals and sensitivity to external influences that offer unique insights.
Genomics is the layer least influenced by environment once a person is born. Of course, mutations can occur and change someone’s DNA, but these happen locally and are most often corrected. In contrast, metabolomics is the layer most sensitive to the environment. It responds to our diet, lifestyle, and exposures. For example, metabolomics profiles can shift in response to year after year of terrible food choices (Limonciel et al. 2013). Poor diet usually leads to chronic low-grade inflammation, which presents metabolic patterns closely associated with markers of inflammation at the granular level (Pietzner et al. 2017).
Genomics can tell you about your risk of developing an inflammatory disease, but it cannot track how this risk evolves throughout your life. Similarly, genomics can identify genotypes that will influence response to a specific drug, but it does not respond to influences from the environment that determine our drug response. Metabolomics can. That’s exactly why it’s such a great tool for population-based medicine where the need for measures of disease risk beyond genetic predisposition is high.
Learn more about the power of multiomics for 5P medicine in my webinar.
Population-based medicine with metabolomics
Population-based medicine takes advantage of the statistical power of large epidemiological cohorts to study health and disease at a different scale. At first glance, this seems like the opposite of precision medicine, but these large cohort studies can be a great source of biomarkers and complex risk scores to evaluate the health of a single individual.
Applying metabolomics in population-based cohort studies provides a wealth of information that enables:
- Metabolic risk scoring
- Metabotyping
- Multiomics integration
Metabolic risk scores
Population-based medicine was among the first disciplines to make use of genomics, designing genome-wide association studies (GWAS) that have identified numerous gene variants associated with the risk of developing disease, from type 2 diabetes (Grant et al. 2006) to coronary artery disease (Helgadottir et al. 2007). However, these gene variants rarely explain more than 10% of the risk for developing complex chronic diseases.
In complex diseases such as Alzheimer’s disease, breast cancer or type 2 diabetes, environmental factors play a massive role, reflected in the low percentage of variance explained by genetic variants (So et al. 2011). This means that much of the remaining 90% is likely to be explained by metabolomics, an omic strongly influenced by the environment.
Understanding the external, environmental factors that associate with a disease or trait enables better risk assessment and prevention, more sensitive diagnosis, and ultimately enables precision medicine for the parts of the population exposed to the external factors identified.
For example, (Hoffman et al. 2024) created a metabolomic risk score that reflects the response of the organism to traffic-related air pollution. To strengthen their model, the authors combined metabolomic data from several cohorts together with simulated data. The power of the associations was relatively low, although the metabolites selected by the models made mechanistic sense, comforting the authors on their relevance to the risk score. The authors emphasized the need for large sample sizes and standardized metabolomics methods to support this approach.
Metabotyping
I’ve discussed this approach in previous blogs in this series, but metabotyping is also a great tool for population-based medicine. Metabotyping consists in forming sub-groups of a population based on their metabolomic profile. This type of clustering is a very good way to uncover sub-phenotypes of a disease that may benefit from distinct early diagnostic tools and different treatments.
On a recent episode of The Metabolomist podcast, Rachel Kelly explains how she and her group have applied this approach in epidemiological studies focused on asthma (Kelly et al. 2021) and autism spectrum disorder (Prince et al. 2023). In both cases, metabolomics was a powerful tool to better understand the condition and begin to think of novel approaches for precision medicine using these cohort-based results.
In 2018, Lacruz et al. showed that not only is metabolomics a great tool to stratify a population through metabotyping, but also that monitoring these metabotypes can be very informative. In the Cardiovascular Disease, Living and Ageing in Halle (CARLA) cohort (n=1,409), nearly 60% of the population had a stable metabotype after a four-year interval. Metabotype instability, however, was associated with a higher risk of all-cause mortality. The authors highlighted metabotype variability as a potential early indicator of pre-clinical disease. You can also listen to Gabi Kastenmüller, the senior author on this publication, discussing the use of metabolomics in this and other studies on large cohorts on the podcast.
Multiomics integration
As mentioned above, the large cohorts used in population-based medicine require robust omics methods to generate meaningful associations. Combining omics is also a way to highlight associations that wouldn’t be visible with a single omics layer.
A well-known example is the integration of metabolomics with GWAS, referred to as mGWAS. I’ve discussed this approach on the podcast with one of the pioneers of mGWAS, Karsten Suhre. In that episode, he explains how using metabolomics and ratios of metabolites helps us identify variants that were previously out of scope, and with excellent p values.
Read more about how to perform mGWAS here.
Large biobanks increasingly include the measurement of multiple omics to enable multiomics analysis for their users. For instance, the Tohoku Medical Megabank Project in Japan created the jMorp database, which compiles multiomics data including genomics, proteomics and metabolomics, as well as related clinical data (Ogishima et al. 2021). These initiatives help to democratize the use of omics in epidemiology and to the development of FAIR science.
Outlook
As the most comprehensive measure of phenotype and external influences on human biology, metabolomics is uniquely positioned to provide the missing pieces of the multiomics puzzle so greatly needed in epidemiology.
From the calculation of metabolic risk scores that can complement or even merge with polygenetic risk scores from genomic studies, to the enhanced understanding of disease provided by metabotyping, metabolomics has a lot to offer to population-base medicine.
We explored the application of this omic to cohort studies in our whitepaper on “FAIR compliant metabolomics profiling of population-based studies.” In particular, we discussed how the level of standardization required for the integration of metabolomics in large cohort studies is a critical point that requires dedicated solutions.
The increasing adoption of metabolomics in large cohort studies and biobanks holds great promise for uncovering the variance related to complex diseases and advancing population-based medicine.
Sign up for our newsletter to be notified when the next blog on 5P medicine comes out.
References
Grant et al.: Variant of transcription factor 7-like 2 (TCF7L2) gene confers risk of type 2 diabetes (2006) Nature Genetics | https://doi.org/10.1038/ng1732
Helgadottir et al.: A common variant on chromosome 9p21 affects the risk of myocardial infarction (2007) Science | https://doi.org/10.1126/science.1142842
Hoffman et al.: Development of a metabolomic risk score for exposure to traffic-related air pollution: A multi-cohort study (2024) Environmental Research | https://doi.org/10.1016/j.envres.2024.120172
Kelly et al.: Metabo-endotypes of asthma reveal differences in lung function: Discovery and validation in two TOPMed cohorts (2021) ATS | http://doi.org/10.1164/rccm.202105-1268OC
Lacruz et al.: Instability of personal human metabotype is linked to all-cause mortality (2018) Nature | https://doi.org/10.1038/s41598-018-27958-1
Limonciel et al.: Complex chronic diseases have a common origin (2013) I https://biocrates.com/wp-content/uploads/2024/07/biocrates-Complex-chronic-diseases-have-a-common-origin.pdf
Ogishima et al.: dbTMM: an integrated database of large-scale cohort, genome and clinical data for the Tohoku Medical Megabank Project (2021) Human Genome Variation | https://doi.org/10.1038/s41439-021-00175-5
Pietzner et al.: Plasma proteome and metabolome characterization of an experimental human thyrotoxicosis model (2017) BMC Medicine | http://doi.org/10.1186/s12916-016-0770-8
Prince et al.: Phenotypically driven subgroups of ASD display distinct metabolomic profiles (2023) Brain, Behavior, and Immunity | https://doi.org/10.1016/j.bbi.2023.03.026
So et al.: Evaluating the heritability explained by known susceptibility variants: a survey of ten complex diseases (2011) Genetic Epidemiology | https://doi.org/10.1002/gepi.20579